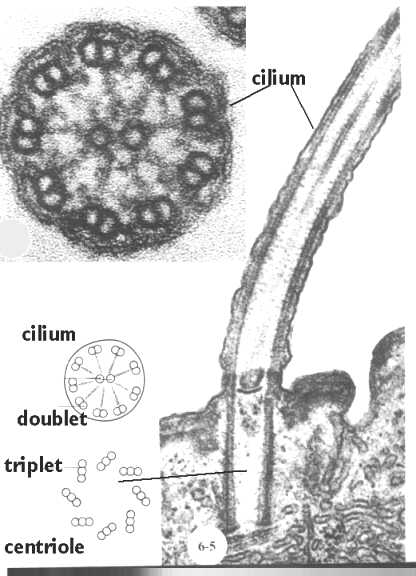
Cilia and flagella are projections from the cell. They are made up of microtubules , as shown in this cartoon. They are motile and designed either to move the cell itself or to move substances over or around the cell. The primary purpose of cilia in mammalian cells is to move fluid, mucous, or cells over their surface. Cilia and flagella have the same internal structure. The major difference is in their length.
This figure shows a cross section of a cilium next to a longitudinal section. Below, we will see how the microtubules are organized in the core (shown in the cartoon in this figure). Also, shown is the
How do Cilia and Flagella move?
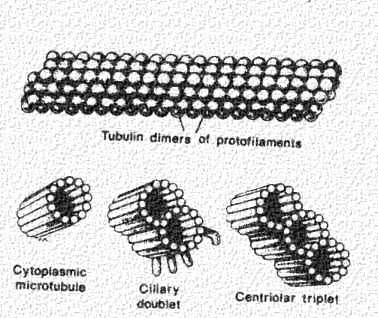
Cilia and flagella move because of the interactions of a set of microtubules inside. Collectively, these are called an "axoneme", This figure shows a microtubule (top panel) in surface view and in cross section (lower left hand panel). Two of these microtubules join to form one doublet in the cilia or flagella This is shown in the middle panel. Note that one of the tubules is incomplete. Furthermore, there are important microtubule associated proteins (MAPs) projecting from one of the microtubule subunits.
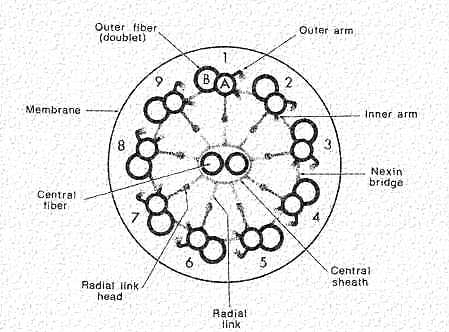
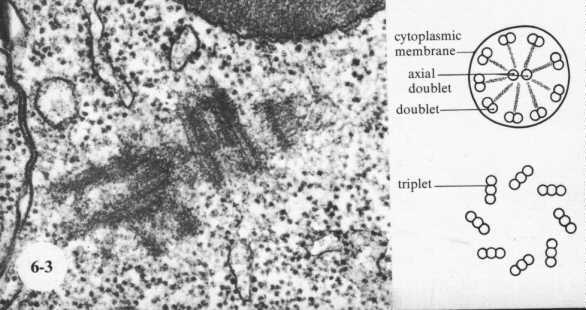
A cross section of a cilium is shown in this cartoon. Note that there is a circle of nine doublets, each of which have one complete (A Tubule) and one incomplete (B Tubule) microtubules. The Core doublets are both complete. Extending from the doublets are sets of arms that join neighboring doublets. These are composed of the protein "dynein". It is spaced at 24 nm intervals. Nexin links are spaced along the microtubules to hold them together. Projecting inward are radial spokes that connect with a sheath enclosing the doublets.
This figure shows an electron micrograph of a cross section of a cilium. Note that you can see the dynein arms and the nexin links. The dynein arms have ATPase activity. In the presence of ATP, they can move from one tubulin to another. They enable the tubules to slide along one another so the cilium can bend. The dynein bridges are regulated so that sliding leads to synchronized bending. Because of the nexin and radial spokes, the doublets are held in place so sliding is limited lengthwise. If nexin and the radial spokes are subjected to enzyme digestion, and exposed to ATP, the doublets will continue to slide and telescope up to 9X their length.
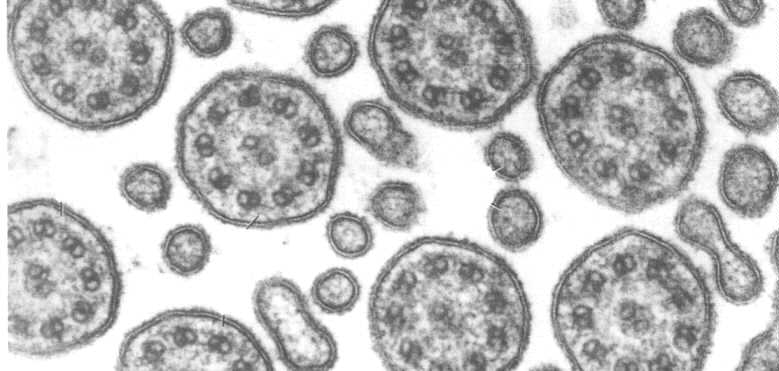
Another micrograph of the cell surface showing a number of cilia. These must be organized functionally so the cilia beat in a wave.
Centrioles and Basal Bodies
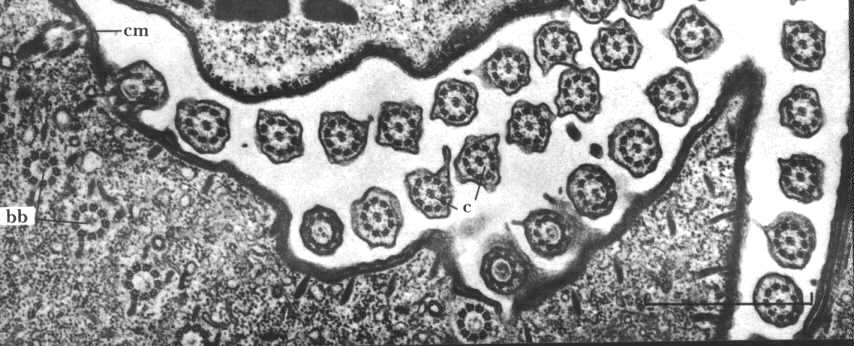
Cilia and flagella are organized from centrioles that move to the cell periphery. These are called "basal bodies" and are shown in this electron micrograph (bb). Note the numerous cilia projecting from the cell membrane (cm). Basal bodies control the direction of movement of the cilia. This can be shown experimentally.
Centrioles control the direction of cilia or flagella movement.
Note the experiment in your text that describes how the direction of
movement was changed. Paramecium has parallel rows of cilia all aligned so that they will
beat in the same direction. However, in the 1960's rows of cilia/basal bodies were grafted
into Paramecium and they were able to show a change in direction of the beat. The cells
passed on the change to future generations even though this was not a genetic alteration.
See you text, page 820, for more information.
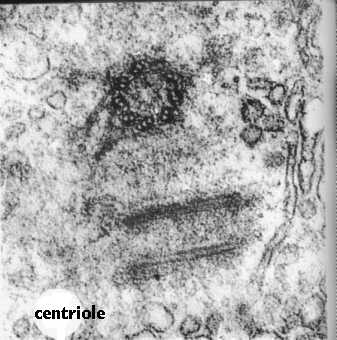
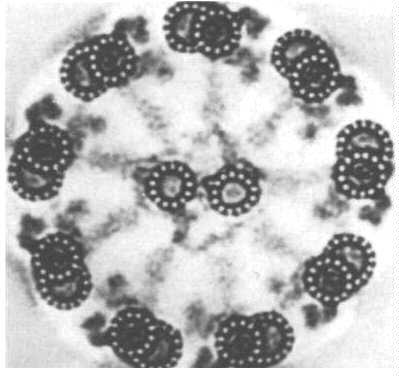
Like Cilia and Flagella, Centrioles are also made of microtubules. The difference is that they contain 9 sets of triplets and no doublet in the center. How the triplets in the basal body turn into the cilium doublet remains a mystery.Centrioles come in pairs, each organized at right angles to the other. This figure shows an electron micrograph of a pair of centrioles and the cartoon compares the cross section of a cilium with that of a centriole. Centrioles organize the spindle apparatus on which the chromosomes move during mitosis. You will learn more about that in the lectures designed to cover this phenomenon.
Central Replication.
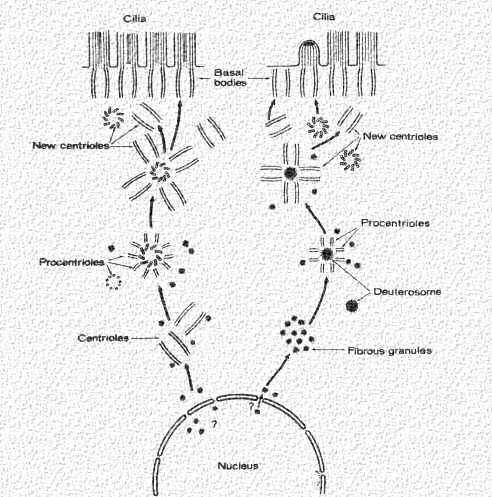
Centrioles replicate autonomously like mitochondria and peroxisomes. They begin from centers which contain proteins needed for their formation (tubulin, etc.), Then the procentrioles form. Each grows out a single microtubule from which the triplet can form. Once a centriole is made, daughter centrioles can grow out from the tubules at right angles as shown in this cartoon. These then add to the daughter cell (in a dividing cell), or they move to the periphery and form the basal body for the cilium.