Lipid Catabolism: Fatty Acids &
Triacylglycerols
|
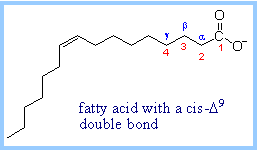
A
16-carbon fatty acid,
with numbering conventions, is shown at right. Most naturally occurring
fatty acids have an even number of carbon atoms. The pathway for catabolism
of fatty acids is referred to as the b-Oxidation
Pathway, because oxidation occurs at the b-carbon
(C3).
|
|
|
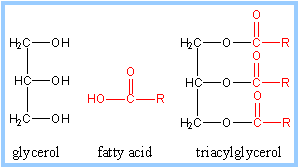
Triacylglycerols
(triglycerides) are the most abundant dietary lipids. They are the form in
which we store reduced carbon for energy. Each triacylglycerol has a
glycerol backbone to which are esterified 3 fatty acids. Most
triacylglycerols are "mixed." The three fatty acids differ in chain length
and number of double bonds
Lipases hydrolyze triacylglycerols, releasing one fatty acid at a time,
producing diacylglycerols, and eventually glycerol.
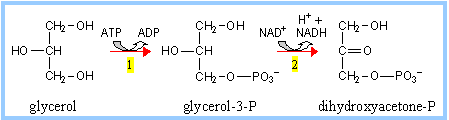
|
|
|
Glycerol
arising from hydrolysis of triacylglycerols is converted to the
Glycolysis
intermediate dihydroxyacetone phosphate, by reactions catalyzed by:
(1) Glycerol Kinase
(2) Glycerol Phosphate Dehydrogenase |
|
Free fatty acids, which in solution
have detergent properties, are transported in the blood bound to albumin, a
serum protein produced by the liver.
Several proteins have been identified that facilitate transport of long chain
fatty acids into cells, including the plasma membrane protein CD36.
|
Fatty
acid activation:
Acyl-CoA Synthases (Thiokinases),
associated with endoplasmic reticulum membranes and the outer mitochondrial
membrane, catalyze activation of long chain fatty acids, esterifying them to
coenzyme A, as shown at right. This process is ATP-dependent, and occurs in
2 steps. There are different Acyl-CoA Synthases for fatty acids of different
chain lengths.
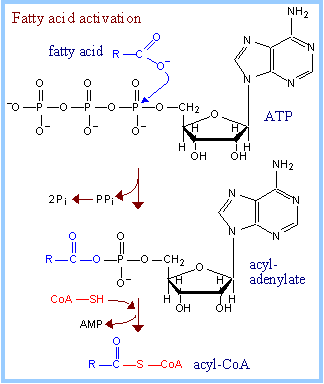
Exergonic hydrolysis of PPi
(P~P), catalyzed by Pyrophosphatase,
makes the coupled reaction spontaneous. Overall,
two
~P
bonds of ATP are
cleaved during fatty acid activation. The acyl-coenzyme A product includes
one "high energy" thioester linkage. |
|
Summary of fatty acid activation:
- fatty acid + ATP à
acyl-adenylate + PPi
PPi à 2 Pi
- acyladenylate + HS-CoA
à acyl-CoA + AMP
Overall: fatty acid + ATP +
HS-CoA à acyl-CoA + AMP + 2 Pi
|
For
most steps of the b-Oxidation
Pathway, there are multiple enzymes specific for particular fatty acid chain
lengths.
Fatty acid
b-oxidation is considered to occur in the mitochondrial
matrix. Fatty acids must enter the matrix to be oxidized.
However enzymes of the pathway specific for very long chain fatty acids are
associated with the inner mitochondrial membrane, facing the matrix.
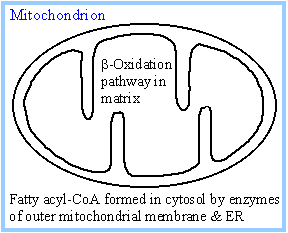
Fatty acyl-CoA formed outside the
mitochondria can pass through the outer mitochondrial membrane, which
contains large VDAC channels, but cannot penetrate the mitochondrial inner
membrane. |
|
|
Transfer of the fatty acid
moiety across the inner mitochondrial membrane involves carnitine. Carnitine
Palmitoyl Transferases catalyze transfer of a fatty acid between the thiol
of Coenzyme A and the hydroxyl on carnitine.
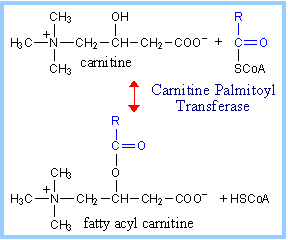
Carnitine-mediated transfer of the fatty acyl
moiety into the mitochondrial matrix is a 3-step process, as presented
below. |
|
|
1.
Carnitine Palmitoyl Transferase I, an enzyme associated with
the cytosolic surface of the outer mitochondrial membrane, catalyzes
transfer of a fatty acid from ester linkage with the thiol of coenzyme A to
the hydroxyl on carnitine.
2. Carnitine Acyltransferase, an
antiporter in the inner
mitochondrial membrane, mediates transmembrane exchange of fatty
acyl-carnitine for carnitine.
3. Within the mitochondrial matrix (or
associated with the matrix surface of the inner mitochondrial membrane,
Carnitine Palmitoyl Transferase II
catalyzes transfer of the fatty acid from carnitine to coenzyme A. (Carnitine
exits the matrix in step 2.) The fatty acid is now esterified to coenzyme A
within the mitochondrial matrix.
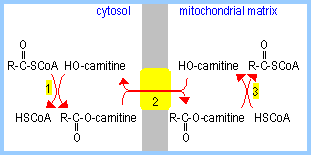
|
|
Control of fatty acid
oxidation is exerted mainly at the step of fatty acid entry into mitochondria.
|
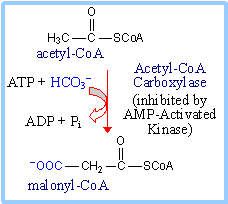
Malonyl-CoA (which is also
a precursor for
fatty acid synthesis)
inhibits Carnitine Palmitoyl Transferase I.
Malonyl-CoA is produced from acetyl-CoA by
the enzyme Acetyl-CoA Carboxylase.
AMP-Activated Kinase a sensor of cellular
energy levels, is allosterically activated by AMP, which is relatively high
in concentration when [ATP] is low.
Acetyl-CoA Carboxylase is inhibited when
phosphorylated by AMP-Activated Kinase, leading to decreased production of
malonyl-CoA.
The decrease in malonyl-CoA concentration
leads to increased activity of Carnitine Palmitoyl Transferase I.
The resulting increased fatty acid oxidation
generates acetyl-CoA, for entry into Krebs cycle with associated ATP
production.
|
|
AMP-Activated Kinase functions under a variety of
conditions that lead to depletion of cellular ATP (reflected as increased AMP),
including glucose deprivation, exercise, hypoxia and ischaemia.
- AMP-Activated Kinase
regulates various metabolic pathways to promote catabolism leading to ATP
synthesis (e.g., stimulation of fatty acid oxidation), while inhibiting
energy-utilizing anabolic pathways (e.g.,
fatty acid synthesis).
- AMP-Activated Kinase in the
hypothalamus of the brain is involved also in regulation of food intake.
b-Oxidation
Pathway:
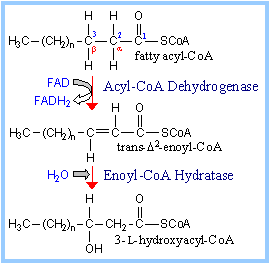
Step 1.
Acyl-CoA Dehydrogenase catalyzes oxidation of the fatty acid moiety of acyl-CoA,
to produce a double bond between carbon atoms 2 and 3.
|
There are different Acyl-CoA
Dehydrogenases for short (4-6 C), medium (6-10 C), long and very long (12-18
C) chain fatty acids. Very Long Chain Acyl-CoA Dehydrogenase is bound to the
inner mitochondrial membrane. The others are soluble enzymes located in the
mitochondrial matrix.
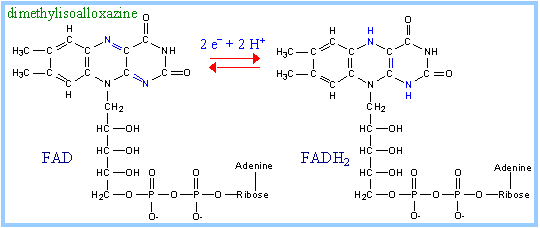
FAD
is the prosthetic group that functions as electron acceptor for Acyl-CoA
Dehydrogenase. Proposed mechanism:
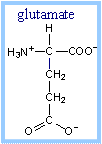
A
glutamate side-chain carboxyl extracts a proton from the
a-carbon of the substrate,
facilitating transfer of 2 e- with H+ (a hydride) from
the b position to FAD. The reduced
FAD accepts a second H+, yielding FADH2.
The carbonyl oxygen of the thioester
substrate is hydrogen bonded to the 2'-OH of the ribityl moiety of FAD,
giving this part of FAD a role in positioning the substrate and increasing
acidity of the substrate a-proton.
The reactive glutamate and FAD are on
opposite sides of the substrate at the active site. Thus the reaction is
stereospecific, yielding a trans double bond in enoyl-CoA.
|
|
|
FADH2
of Acyl CoA Dehydrogenase is reoxidized by transfer of 2 electrons to an
Electron Transfer Flavoprotein (ETF),
which in turn passes the electrons to coenzyme Q of the respiratory chain.
|
|
Step 2. Enoyl-CoA Hydratase catalyzes
stereospecific hydration of the trans double bond produced in the 1st step of
the pathway, yielding L-hydroxyacyl-Coenzyme A .
|
Step 3. Hydroxyacyl-CoA
Dehydrogenase catalyzes oxidation of the hydroxyl in the
b position (C3) to a ketone. NAD+
is the electron acceptor.
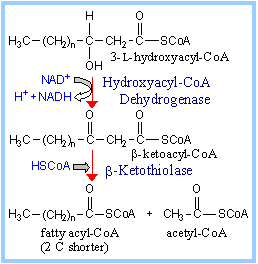
Step 4.
b-Ketothiolase (b-Ketoacyl-CoA
Thiolase) catalyzes thiolytic cleavage.
Proposed mechanism: A cysteine S attacks the
b-keto C. Acetyl-CoA is released,
leaving the fatty acyl moiety in thioester linkage to the cysteine thiol.
The thiol of HSCoA displaces the cysteine thiol, yielding fatty acyl-CoA (2
C shorter).
A membrane-bound trifunctional protein
complex with two subunit types expresses the enzyme activities for steps 2-4
of the b-oxidation pathway for long
chain fatty acids. Equivalent enzymes for medium and short chain length
fatty acids are soluble proteins of the mitochondrial matrix. |
|
Summary of
one round of the b-oxidation pathway:
fatty acyl-CoA + FAD + NAD+
+ HS-CoA à
fatty acyl-CoA (2 C shorter) + FADH2 + NADH + H+
+ acetyl-CoA
The b-oxidation
pathway is cyclic. The
product, 2 carbons shorter, is the input to another round of the pathway. If, as
is usually the case, the fatty acid contains an even number of C atoms, in the
final reaction cycle butyryl-CoA is converted to 2 copies of acetyl-CoA.
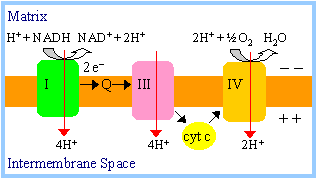
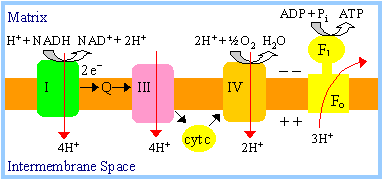
ATP production:
-
NADH produced during
fatty acid oxidation is reoxidized by transfer of 2e-
to respiratory chain complex I. Transfer of 2e-
from complex I to oxygen causes sufficient proton ejection to yield
approximately 2.5 ATP.
(Recall that 4H+ enter the mitochondrial matrix per ATP
synthesized, taking into account transmembrane flux of ADP, ATP & Pi;
see notes on
oxidative
phosphorylation).
-
FADH2 of Acyl
CoA Dehydrogenase is reoxidized by transfer of 2e-
via ETF to
coenzyme Q of the respiratory chain. H+ ejection from the
mitochondrial matrix that accompanies transfer of 2e-
from coenzyme Q to oxygen leads to production of approximately
1.5 ATP.
-
Acetyl-CoA can enter
Krebs cycle,
where the acetate is oxidized to CO2, yielding additional NADH,
FADH2, and ATP.
|
|
Fatty acid oxidation is a major source of cellular ATP.
The reactions presented above accomplish catabolism of a fatty acid
with an even number of carbon atoms and no double bonds. Additional enzymes deal
with catabolism of fatty acids with an odd number of carbon atoms or including
double bonds.
- The final round of
b-oxidation of a fatty acid with an
odd number of carbon atoms yields acetyl-CoA and
propionyl-CoA. Propionyl-CoA
is converted to the Krebs cycle intermediate succinyl-CoA, by a pathway
involving vitamin B12. That pathway is discussed along with the
topic of amino acid catabolism. Catabolism of some amino acids also yields
propionyl-CoA.
- Most double bonds of
naturally occurring fatty acids have the cis configuration. As carbon atoms
are removed two at a time, a double bond may end up in the wrong position or
wrong configuration to be the correct substrate for Enoyl-CoA Hydratase. The
reactions that allow unsaturated fatty acids to be fully catabolized by the
b-oxidation pathway.
|
Peroxisomal Fatty
oxidation:
b-Oxidation of very long chain
fatty acids also occurs within peroxisomes. FAD
is electron acceptor for peroxisomal
Acyl-CoA Oxidase, which
catalyzes the first oxidative step of the pathway. The resulting FADH2
is reoxidized in the peroxisome producing hydrogen peroxide:
FADH2 + O2
à
FAD + H2O2
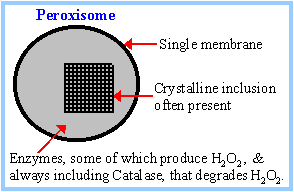
The peroxisomal enzyme Catalase degrades H2O2
by the reaction:
2
H2O2
à
2
H2O + O2
These reactions produce
no ATP. Once fatty acids
are reduced in length within the peroxisomes they may shift to the
mitochondria to be catabolized all the way to CO2. Carnitine is
also involved in transfer of fatty acids into and out of peroxisomes.
|
|
|
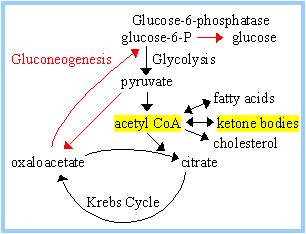
KETONE BODIES:
During fasting or carbohydrate
starvation, oxaloacetate is depleted in liver because it is used for
gluconeogenesis. This impedes entry of acetyl-CoA into Krebs cycle.
Acetyl-CoA then is converted in liver mitochondria to ketone bodies,
acetoacetate and
b-hydroxybutyrate.
|
|
|
Three enzymes are involved
in synthesis of ketone bodies:
b-Ketothiolase.
The final step of the b-oxidation
pathway runs backwards, condensing 2 acetyl-CoA to produce acetoacetyl-CoA,
with release of one CoA.
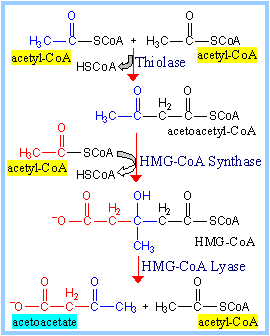
HMG-CoA Synthase catalyzes condensation of a
third acetate moiety (from acetyl-CoA) with acetoacetyl-CoA to form
hydroxymethylglutaryl-CoA (HMG-CoA).
HMG-CoA Lyase cleaves HMG-CoA
to yield acetoacetate plus acetyl-CoA. |
|
|
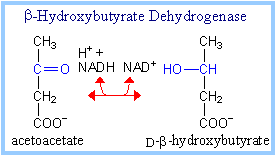
b-Hydroxybutyrate Dehydrogenase
catalyzes inter-conversion of the ketone bodies acetoacetate and
b-hydroxybutyrate.
Ketone bodies are transported in the blood to
other cells, where they are converted back to acetyl-CoA (diagram p. 929)
for catabolism in Krebs cycle, to generate ATP. While ketone bodies thus
function as an alternative fuel, amino acids must be degraded to supply
input to gluconeogenesis when hypoglycemia occurs, since acetate cannot be
converted to glucose. |
|