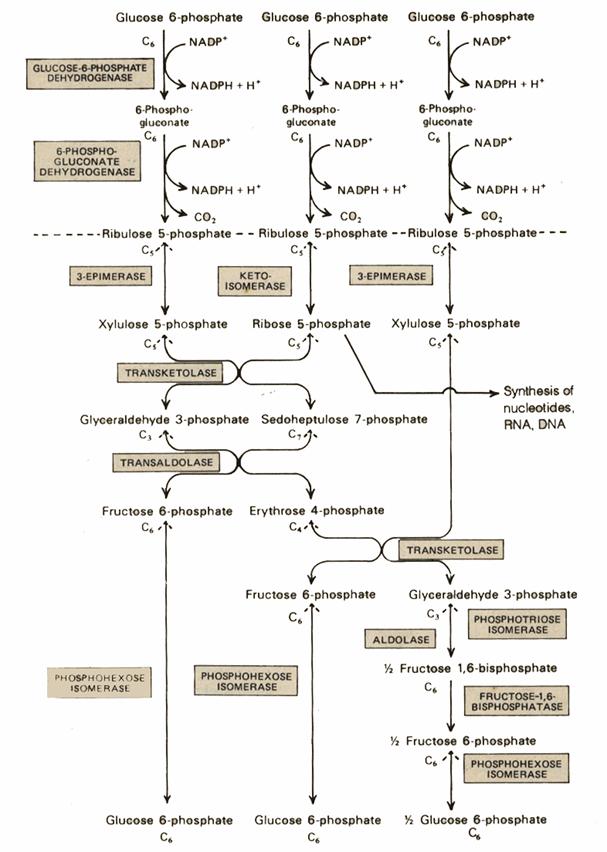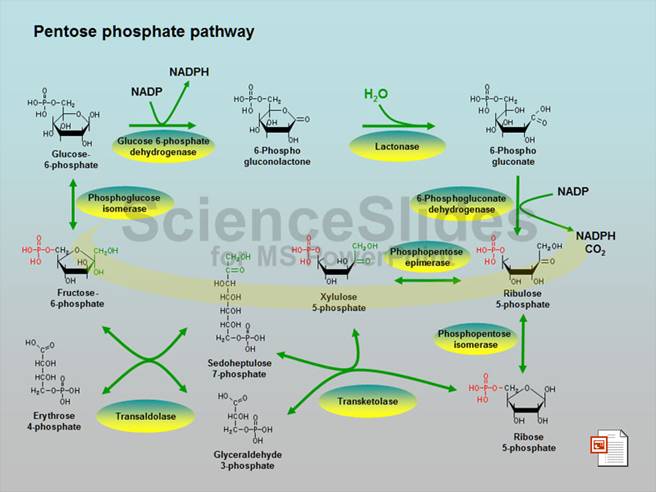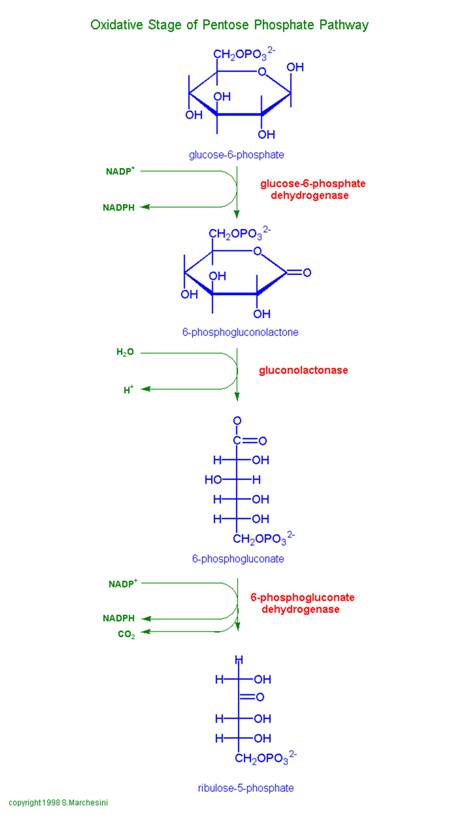
This is also called as Phosphogluconate Pathway, or Hexose Monophosphate Shunt. The linear portion of the pathway carries out oxidation and decarboxylation of glucose-6-phosphate, producing the 5-C sugar ribulose-5-phosphate.
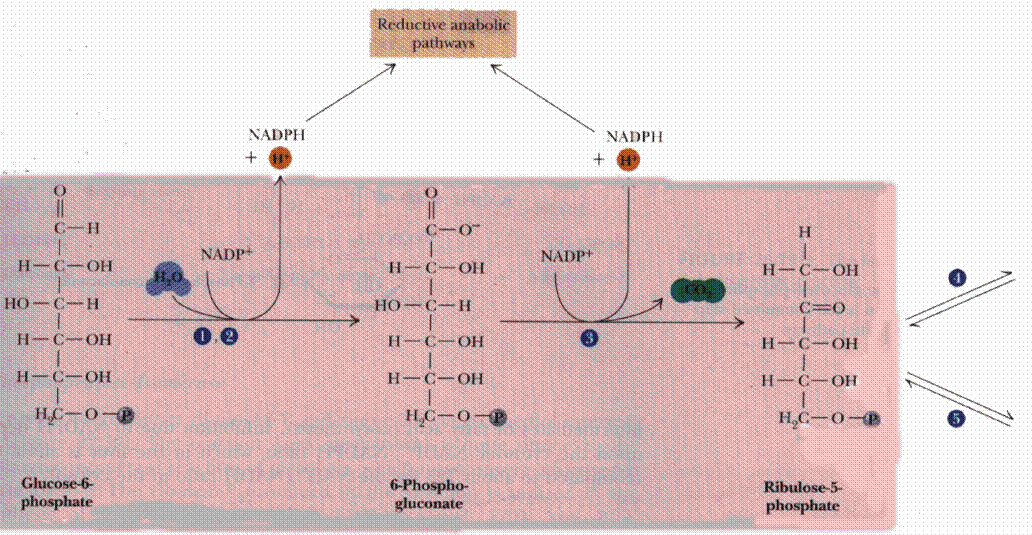
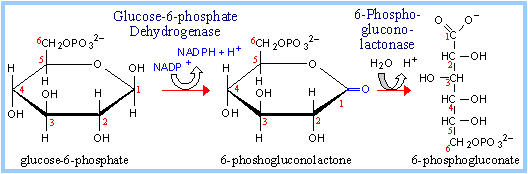
Glucose-6-phosphate Dehydrogenase catalyzes oxidation of the aldehyde (hemiacetal), at C1 of glucose-6-phosphate, to a carboxylic acid in ester linkage (lactone). NADP+ serves as electron acceptor.
6-Phosphogluconolactonase catalyzes hydrolysis of the ester linkage (lactone) resulting in ring opening. The product is 6-phosphogluconate. Although ring opening occurs in the absence of a catalyst, 6-Phosphogluconolactonase speeds up the reaction, decreasing the lifetime of the highly reactive, and thus potentially toxic, 6-phosphogluconolactone.
|
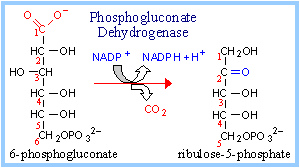
Phosphogluconate Dehydrogenase catalyzes oxidative decarboxylation of 6-phosphogluconate, to yield the 5-C ketose ribulose-5-phosphate. The hydroxyl at C3 (C2 of the product) is oxidized to a ketone. This promotes loss of the carboxyl at C1 as CO2. NADP+ again serves as oxidant (electron acceptor).
|
|
|
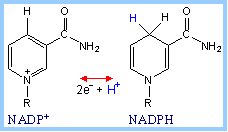
Reduction of NADP+ (as with NAD+) involves transfer of 2e- plus 1H+ to the nicotinamide moiety.
|
|
|
NAD+ and NADP+ differ only in the presence of an extra phosphate on the adenosine ribose of NADP+. This difference has little to do with redox activity, but is recognized by substrate-binding sites of enzymes. It is a mechanism for separation of catabolic and synthetic pathways.
NADPH, a product of the Pentose Phosphate Pathway, functions as a reductant in various synthetic (anabolic) pathways, including fatty acid synthesis. NAD+ serves as electron acceptor in catabolic pathways in which metabolites are oxidized. The resultant NADH is reoxidized by the respiratory chain, producing ATP. |
|
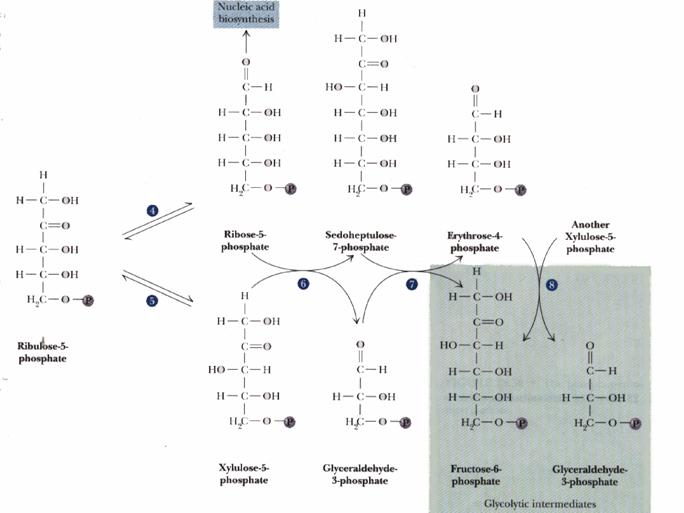
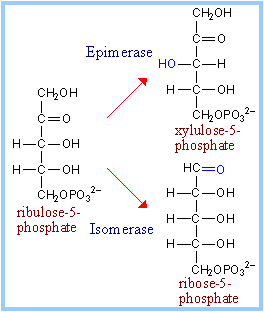
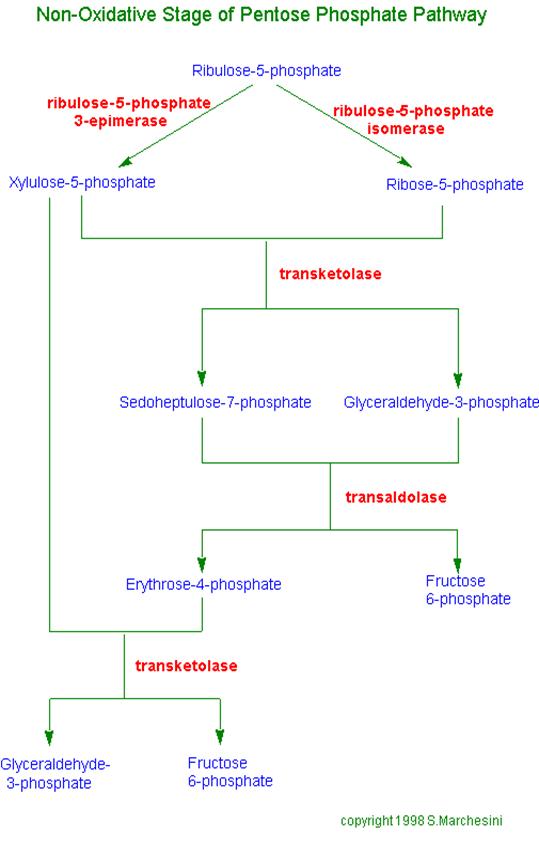
The remainder of the Pentose Phosphate Pathway accomplishes conversion of the 5-C ribulose-5-phosphate to the 5-C product ribose-5-phosphate, or to the 3-C glyceraldehyde-3-phosphate and the 6-C fructose-6-phosphate (reactions 4 to 8 p. 863).
|
Additional enzymes include Ribulose-5-phosphate Epimerase, Ribulose-5-phosphate Isomerase, Transketolase, and Transaldolase.
|
|
Transketolase and Transaldolase catalyze transfer of 2-C and 3-C molecular fragments respectively, in each case from a ketose donor to an aldose acceptor. D. E. Nicholson has suggested that the names of these enzymes should be changed, since Transketolase actually transfers an aldol moiety (glycoaldehyde) and Transaldolase actually transfers a ketol moiety (dihydroxyacetone). However the traditional enzyme names are used here.
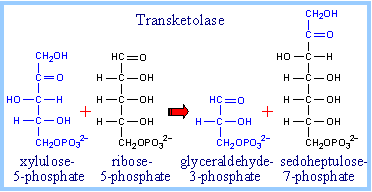
|
Transketolase transfers a 2-C fragment from xylulose-5-phosphate to either ribose-5-phosphate or erythrose-4-phosphate. |
|
|
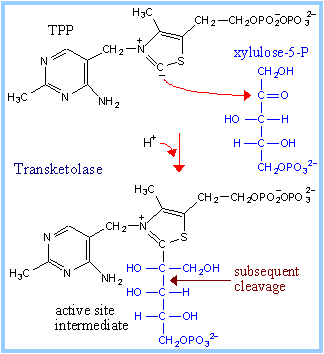
Transketolase utilizes as prosthetic group thiamine pyrophosphate (TPP), a derivative of vitamin B1. H+ readily dissociates from the C between N and S in the thiazolium ring. |
|

TPP binds at the active sites of enzymes in a "V" conformation. The amino group of the aminopyrimidine moiety is close to the dissociable proton, and serves as the proton acceptor. This proton transfer is promoted by a glutamate residue adjacent to the pyrimidine ring. The thiazolium carbanion (ylid) that results from proton dissociation reacts with the carbonyl C of xylulose-5-P to form an addition compound.
|
The positively charged N in the thiazole ring acts as an electron sink, promoting C-C bond cleavage. The 3-C aldose glyceraldehyde-3-phosphate is released. A 2-C fragment remains on TPP. Completion of the reaction is by reversal of these steps. The 2-C fragment condenses with one of the aldoses erythrose-4-phosphate (4-C) or ribose-5-phosphate (5-C) to form a 6-C or 7-C ketose-phosphate product.
Transfer of the 2-C fragment to the 5-C aldose ribose-5-phosphate yields sedoheptulose-7-phosphate (see above). Transfer instead to the 4-C aldose erythrose-4-phosphate yields fructose-6-phosphate. See also diagram p. 865. |
|
|
|
|
|
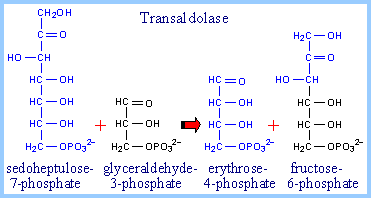
Transaldolase catalyzes transfer of a 3-C dihydroxyacetone moiety, from sedoheptulose-7-phosphate to glyceraldehyde-3-phosphate.
|
|
|
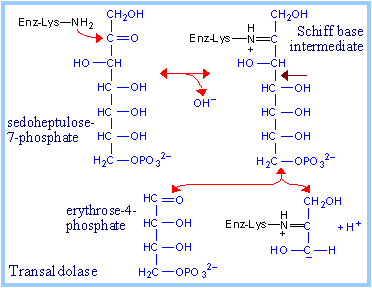
The e-amino group of an active site lysine residue reacts with the carbonyl C of sedoheptulose-7-phosphate to form a protonated Schiff base intermediate. Aldol cleavage results in release of erythrose-4-phosphate. The Schiff base stabilizes the carbanion on C3.
Completion of the reaction occurs by reversal, as the carbanion attacks instead the aldehyde carbon of the 3-carbon aldose glyceraldehyde-3-phosphate to yield the 6-carbon fructose-6-phosphate. |
|
|
|
|
|
The flow of intermediates containing 15 C atoms through Pentose Phosphate Pathway reactions by which 5-C sugars are converted to 3-C and 6-C sugars is summarized in the diagram at right and balance sheet below.
C5 + C5 à C3 + C7 (Transketolase) C3 + C7 à C6 + C4 (Transaldolase) C5 + C4 à C6 + C3 (Transketolase) _______________________________________ 3 C5 à 2C6 + C3 (Overall) Glucose-6-phosphate Dehydrogenase is the committed step of the Pentose Phosphate Pathway. This enzyme is regulated by availability of the substrate NADP+. As NADPH is utilized in reductive synthetic pathways, the increasing concentration of NADP+ stimulates the Pentose Phosphate Pathway, to replenish NADPH. The remainder of the Pentose Phosphate Pathway accomplishes conversion of the 5-C ribulose-5-phosphate to the 5-C product ribose-5-phosphate, or to the 3-C glyceraldehyde-3-phosphate and the 6-C fructose-6-phosphate Glucose-6-phosphate may be regenerated from either the 3-C product glyceraldehyde-3-phosphate or the 6-C product fructose-6-phosphate, via enzymes of Gluconeogenesis. In the diagram at right, IS = Isomerase, EP = Epimerase, TK = Transketolase, TA = Transaldolase. |
|
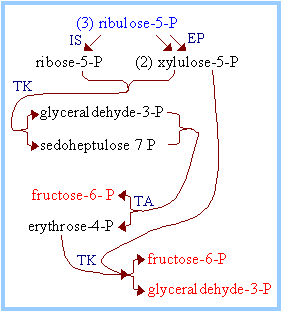
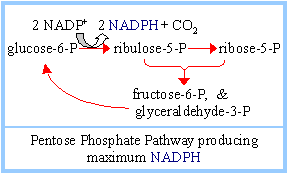
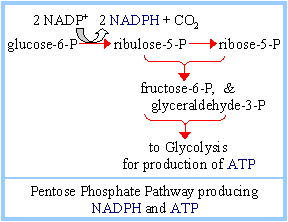
Depending on relative needs of a cell for ribose-5-phosphate, NADPH, and ATP, the Pentose Phosphate Pathway can operate in various modes, to maximize different products. There are three major scenarios:
|
1. Ribulose-5-phosphate may be converted to ribose-5-phosphate, a substrate for synthesis of nucleotides and nucleic acids. The pathway also produces some NADPH.
|
|
|
2. Glyceraldehyde-3-phosphate and fructose-6-phosphate, formed from the 5-carbon sugar phosphates, may be converted to glucose-6-phosphate for reentry into the linear portion of the Pentose Phosphate Pathway, maximizing formation of NADPH.
|
|
|
3. Glyceraldehyde-3-phosphate and fructose-6-phosphate, formed from the 5-carbon sugar phosphates, may enter Glycolysis, for synthesis of ATP. The pathway also produces some NADPH.
Ribose-1-phosphate generated during catabolism of nucleosides also enters the Glycolytic pathway in this way, after first being converted to ribose-5-phosphate. Thus the Pentose Phosphate Pathway serves as an entry into Glycolysis for both 5-carbon and 6-carbon sugars.
|
|