LACOPERON
Operon generally are of two types namely anabolic operon and catabolic operon. Those operon directing the synthesis of enzymes responsible for catabolic reactions then they are known as catabolic operon where as operon coding enzymes for synthetic reactions are known as anabolic operons. Lac operon is an fine example for catabolic operon because it codes for the enzymes which are responsible for degradation of lactose.
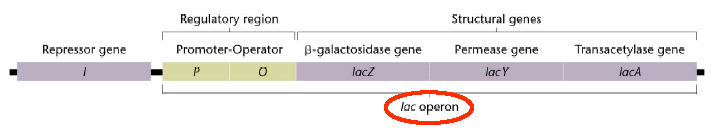
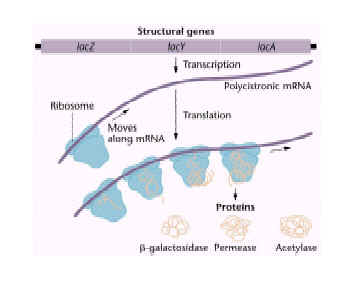
Lac operon consists of lacZ,lacY,lacA genes as structural genes and operator,promoter and suppressor genes as regulatory genes.
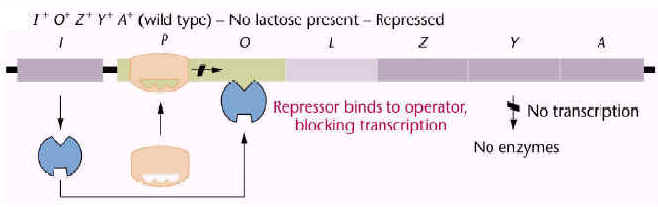
The regulatory gene codes for a regulatory protein. The lac repressor, encoded by the lacI gene, is the regulatory protein of the lac operon.
The operator is the region of DNA of the operon that is the binding site for the regulatory protein.
The promoter is the DNA sequence of the operon recognized by DNA-dependent RNA polymerase. The initiation site for RNA synthesis is immediately downstream of the promoter. The gene for DNA-dependent RNA polymerase is not part of the operon, since the RNA polymerase enzyme transcribes all bacterial operons.
The operon encodes one or more genes for inducible enzymes. The lac operon encodes enzymes necessary for lactose metabolism, including ß-galactosidase, ß-galactoside permease, and ß-galactoside transacetylase
REGULATION:
E. coli prefers to use glucose as an energy source when both glucose and lactose are available. Lactose is an alternative energy source that can be used if glucose is absent. The overall rate of messenger RNA synthesis from the lac operon, and from other operons for alternate catabolic energy sources, is indirectly regulated by the concentration of glucose in the cell.The following table is a summary of the concepts presented in this problem set on the regulation of lac mRNA synthesis by intracellular levels of lactose and glucose. The relative concentration of cAMP and the relative levels of lac mRNA synthesis in the presence or absence of glucose and lactose are shown.
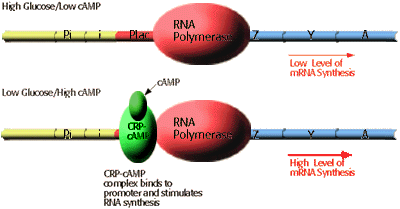
Glucose cAMP Lactose lac mRNA present low concentration absent no mRNA present low concentration present low level of mRNA absent high concentration absent no mRNA absent high concentration present high level of mRNA
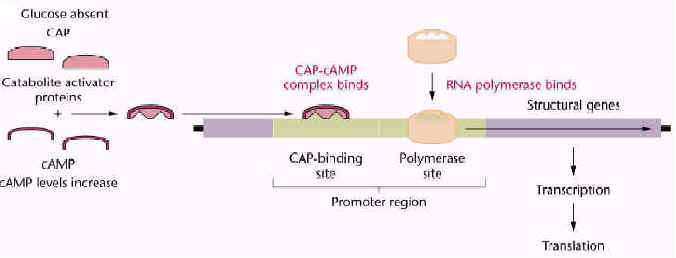
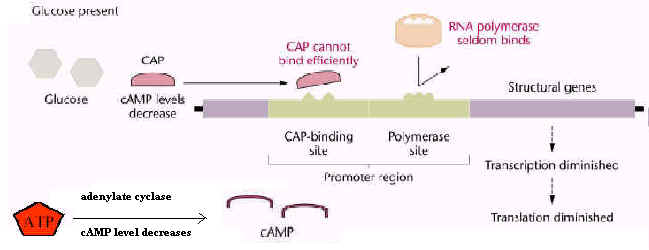
cAMP is the second messenger used directly in the regulation of lac operon expression in response to changing levels of glucose. Control of the rate of transcription of an operons for catabolic enzymes such as the lac operon is regulated through intracellular levels of 3'-5-cyclic adenosine monophosphate, or cAMP. cAMP is synthesized from ATP by the enzyme adenyl cyclase.
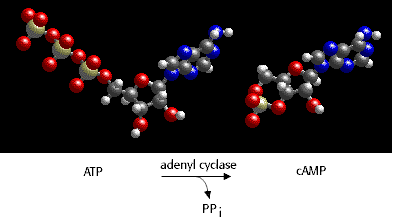
The intracellular level of cAMP is in turn regulated by the intracellular concentration of glucose. cAMP is a co-activator of the CRP protein, a transcriptional enhancer for operons that encode genes for catabolism of energy sources other than glucose, including the lac operon. As glucose levels fall, cAMP levels rise and transcription of an induced lac operon is enhanced. As the levels of glucose rise, cAMP levels fall, and lac operon mRNA synthesis is not enhanced.
mRNA synthesis from the E. coli lac operon is inducible. mRNA synthesis does not occur unless lactose is available for catabolism. The level of mRNA synthesis from the lac operon is also regulated by the intracellular concentration of glucose via a mechanism called catabolite repression.
Briefly, maximum rates of lac mRNA synthesis occur only at low levels of glucose. As suggested by the diagram, cAMP present in cells in response to low glucose levels binds to the CRP protein and the cAMP-CRP complex enhances the level of mRNA synthesis from the lac operon as much as 50 fold. CRP alone cannot activate transcription.
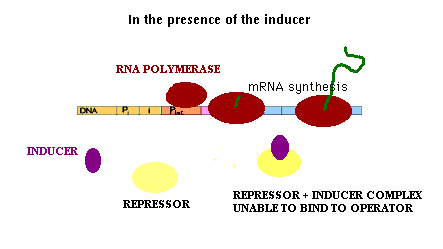
| The inducer is a small molecule that is a signal within the cell that lactose is available for metabolism. The natural inducer is allolactose, an isomer of lactose. Instead, two sulfur-containing analogues of lactose, in particular isopropylthiogalactoside(IPTG) and thiomethylgalactose(TMG) which are effective inducers without being substrates of Beta galactosidase, are used. Inducers having this property are called gratuitous inducers. |
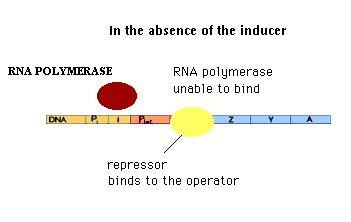
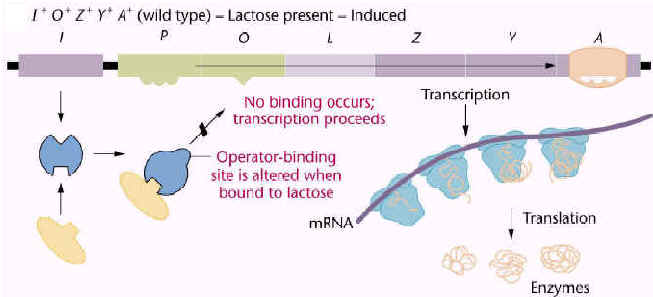
| In the absence of the inducer, the repressor protein binds to the lac operon DNA and prevents synthesis of the mRNA for the lac operon. Since lactose is not available for metabolism, the cell has no need to express enzymes required for the metabolism of lactose. |
|
In the presence of the inducer, the inducer binds to the repressor protein "inducing" a conformational change in the structure of the repressor protein. The repressor protein-inducer complex does not bind to the lac operator DNA. mRNA synthesis from the lac operon can occur when inducer is present.
This regulatory circuit allows E. coli to turn off expression of genes for lactose metabolism when lactose is unavailable, and turn on these genes when lactose is present. |
|
The term lac Z There are several types of mutation that can cause a lac Z Non-sense mutationA codon for an amino acid is mutated to a termination codon (UAA, UAG, UGA), causing premature termination during protein synthesis;Mis-sense mutationA codon for an amino acid is mutated to a codon for a different amino acid. The new amino acid substitution renders the enzyme inactive;Deletion mutationA portion of the DNA of the lac Z gene is absent. With enzyme coding information absent, a functional enzyme cannot be synthesized. |
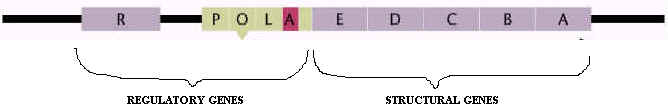
trp operon is an example for anabolic operon. It generally codes for enzymes responsible for the synthesis of tryptophan, an essential aminoacid. trp operon consists of five genes a,b,c,d,e for five enzymes and regulatory genes.
a)trp operon structure:

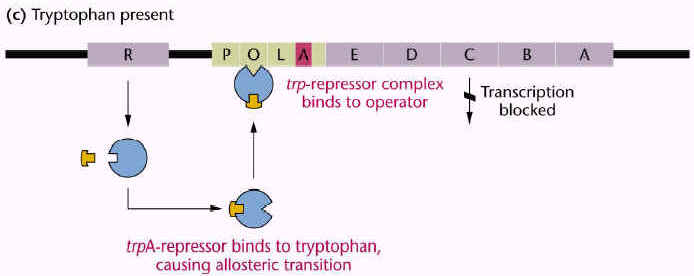
Expression of trp operon was also controlled by an another mechanism known as attenuation.
(refer notes under prokaryotic transcription regulation)